逆境中心朱健康研究组和段成国研究组发现新的蛋白复合体控制异染色质化介导的RNA加工机制
8月29日,PNAS杂志在线发表了中国科学院上海植物逆境生物学研究中心朱健康研究组和段成国研究组题为“A protein complex regulates RNA processing of intronic heterochromatin-containing genes in Arabidopsis”的研究论文。该研究利用生物化学手段鉴定到一个染色质调控因子ASI1的互作蛋白-AIPP1(ASI1 IMMUNOPRECIPITATION PROTEIN 1),发现AIPP1可以作为“桥梁”蛋白介导ASI1和EDM2在细胞内的互作,形成蛋白复合体共同在内含子含有异染色质组分基因的RNA正确加工中起作用。
在真核生物的基因组中,转座子和重复序列(transposable and repetitive elements, TREs)占了很大的比例,其中有很多TRE插入到基因的内含子区,如人类和小鼠基因组中60%的转座子位于内含子中(Sela et al., 2007 Genome Biology),玉米超过10%的内含子含有大于1kb的转座子插入(West et al., 2014 Plos One)。这些内含子TRE经常被DNA甲基化和组蛋白分子标记修饰而异染色质化。通常来说,异染色质化对于临近基因的表达具有抑制效应,有趣的是,我们发现内含子中TRE等异染色质组分的插入并不影响所在基因的正常表达。但是对于其中的机制并不清楚。
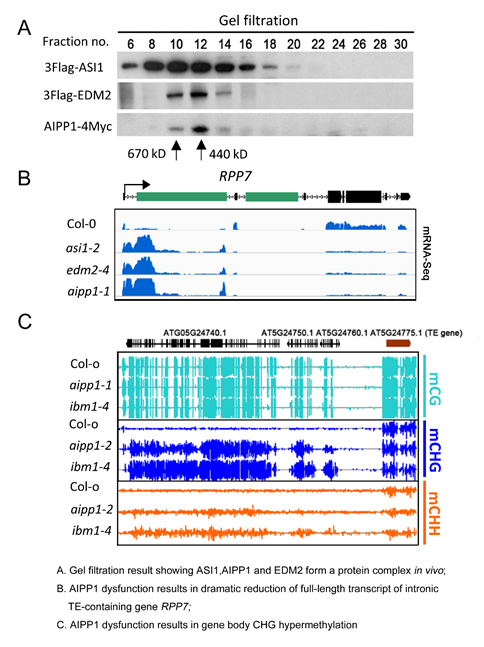
在我们之前的研究中,发现染色质调控因子ASI1 (ANTI-SILENCING 1)和组蛋白H3K9me2结合蛋白EDM2(enhanced downy mildew 2)能够与内含子中的异染色质组分互作控制所在基因在末端进行多聚腺苷酸化(polyA),从而促进全长转录本的产生,即异染色质化介导的RNA加工机制(Wang et al., 2013 PNAS; Lei et al., 2014 PNAS)。虽然ASI1与EDM2功能相似,但是对于二者在RNA加工途径中的互作机制并不清楚。在本研究中,研究人员利用生化手段鉴定到ASI1的潜在互作蛋白:AIPP1,AIPP2,AIPP3以及CPL2。其中AIPP1能够作为一种“桥梁”蛋白介导ASI1和EDM2在细胞内的连接,并形成一个蛋白复合体—AAE复合体(ASI1-AIPP1-EDM2)。与ASI1和EDM2类似,AIPP1功能缺失突变导致大量内含子含有TRE的基因产生大量短转录本,而功能性全长转录本几乎消失,其中包括组蛋白H3K9me2去甲基化酶编码基因IBM1。IBM1能够抑制蛋白编码基因的CHG超甲基化,因此在aipp1突变体中,基因编码区发生CHG高甲基化。更有趣的是,含有PHD结构域的AIPP2,含有BAH结构域的AIPP3和CPL2三者之间也能形成一个复合体,并且其在RNA加工机制中的作用与AAE复合体之间显著不同,说明异染色质介导的RNA加工通路存在着更为复杂的调控机制。该发现对于我们理解高等真核生物中表观遗传修饰介导的异染色质沉默与RNA加工机制之间的互作关系具有重要的意义。