

张鹏研究组揭示植物蓝光受体CRY复合体组装与信号传递的分子机制
2025年12月3日,国际知名学术期刊 Science Advances同日在线发表了中国科学院分子植物科学卓越创新中心张鹏研究员团队与合作单位完成的两项研究成果。研究论文题目分别为“Structural assembly of maize CRY-GL2 photosignaling complex provides insights into its regulatory role in cuticular wax biosynthesis”和“Molecular basis of very-long-chain fatty acid elongation by the CER6-GL2 enzyme complex in plant wax biosynthesis”。这两项工作系统揭示了植物蓝光受体隐花素(CRY)在感知蓝光信号后,如何与下游效应蛋白GLOSSY2(GL2)组装形成信号转导复合体,进而调控植物表皮蜡质合成的分子过程。
光是植物进行光合作用的能量基础,同时也是调控其生长发育的关键环境信号。植物主要通过光受体蛋白感知不同波段的光信号,其中隐花素CRY负责感知350–500 nm波长的蓝光。CRY被蓝光激活后,可与多种下游效应蛋白互作,广泛参与植物的光形态建成、开花时间调控等重要生理过程。张鹏研究组曾于2020年首次解析了玉米与拟南芥中CRY1/CRY2在光激活状态下的四聚体结构,揭示了其光激活机制与分子过程(Shao et al., Nat Struct Mol Biol, 2020;doi: 10.1038/s41594-020-0420-x)。然而,光激活后的CRY如何与下游效应蛋白组装成功能复合体并传递光信号,仍是领域内未解的关键科学问题。
在第一项研究中,团队聚焦玉米CRY如何与蜡质合成关键蛋白GL2(BAHD酰基转移酶家族成员)组装形成有功能的光信号复合体。通过高分辨率晶体结构解析与生化功能分析,研究人员发现蓝光激活后的CRY1能够形成同源四聚体,进而特异性地结合四分子GL2,组装成化学计量比为4:4的异源八聚体光信号复合体(图1A)。研究同时揭示了CRY1四聚体界面重塑后特异性识别GL2的分子基础,并发现这一结合机制在进化上高度保守。该成果首次在原子水平展示了植物CRY光信号复合体的组装过程,为理解光信号转导提供了关键分子依据。该工作由张鹏团队与深圳大学刘宏涛团队、上海师范大学俞芳团队合作完成,张鹏团队博士后刘亚琪与刘宏涛团队副研究员赵志伟为共同第一作者。
在另一项研究中,研究人员聚焦于GL2在植物表皮蜡质合成中的功能机制。在以往的研究中,人们普遍认为GL2作为BAHD家族酰基转移酶,参与植物表皮蜡质主要成分―超长链脂肪酸的延伸/合成过程。本研究中发现,作为BAHD家族酰基转移酶成员的GL2本身并不发挥催化活性,而是与脂肪酸延伸酶CER6(酮酯酰合酶)形成2:2的异源四聚体复合体,从而催化超长链脂肪酸的延伸与合成。结构分析表明,GL2通过N端结构域重塑CER6的底物(酰基辅酶A)结合通道,形成连续疏水腔,从而实现C28至C32超长链脂肪酸的延伸与合成(图1B);并提出CER6可以结合不同的GL2类蛋白,从而实现蜡质成分中不同长度超长链脂肪酸的延伸。张鹏团队博士后刘亚琪与博士生陈媛为该项研究的共同第一作者。
对CRY1c-GL2和CER6-GL2复合体三维结构的进一步分析发现,GL2使用同一界面分别与CRY1c和CER6结合(图1C)。酶活实验表明,CRY1c可剂量依赖性地抑制CER6-GL2复合体的酶活,提示植物可能通过光激活CRY竞争性结合GL2,进而调控蜡质合成(图1D)。尽管其在生物体内的调控功能有待进一步研究,该发现为光调控植物蜡质代谢通路提供了新的分子模型。
综上所述,研究工作系统地揭示了植物体光受体CRY感知环境光信号激活、信号复合物组装到代谢调控的完整信号传导链条(图2)。
本研究获得上海光源、复旦大学、中国科学院生物与化学交叉中心等单位在数据收集方面的支持;研究获国家重点研发计划、国家自然科学基金、中国科学院及上海市科技计划等项目资助。
原文链接:
https://www.science.org/doi/10.1126/sciadv.adz0136
https://www.science.org/doi/10.1126/sciadv.adz0135
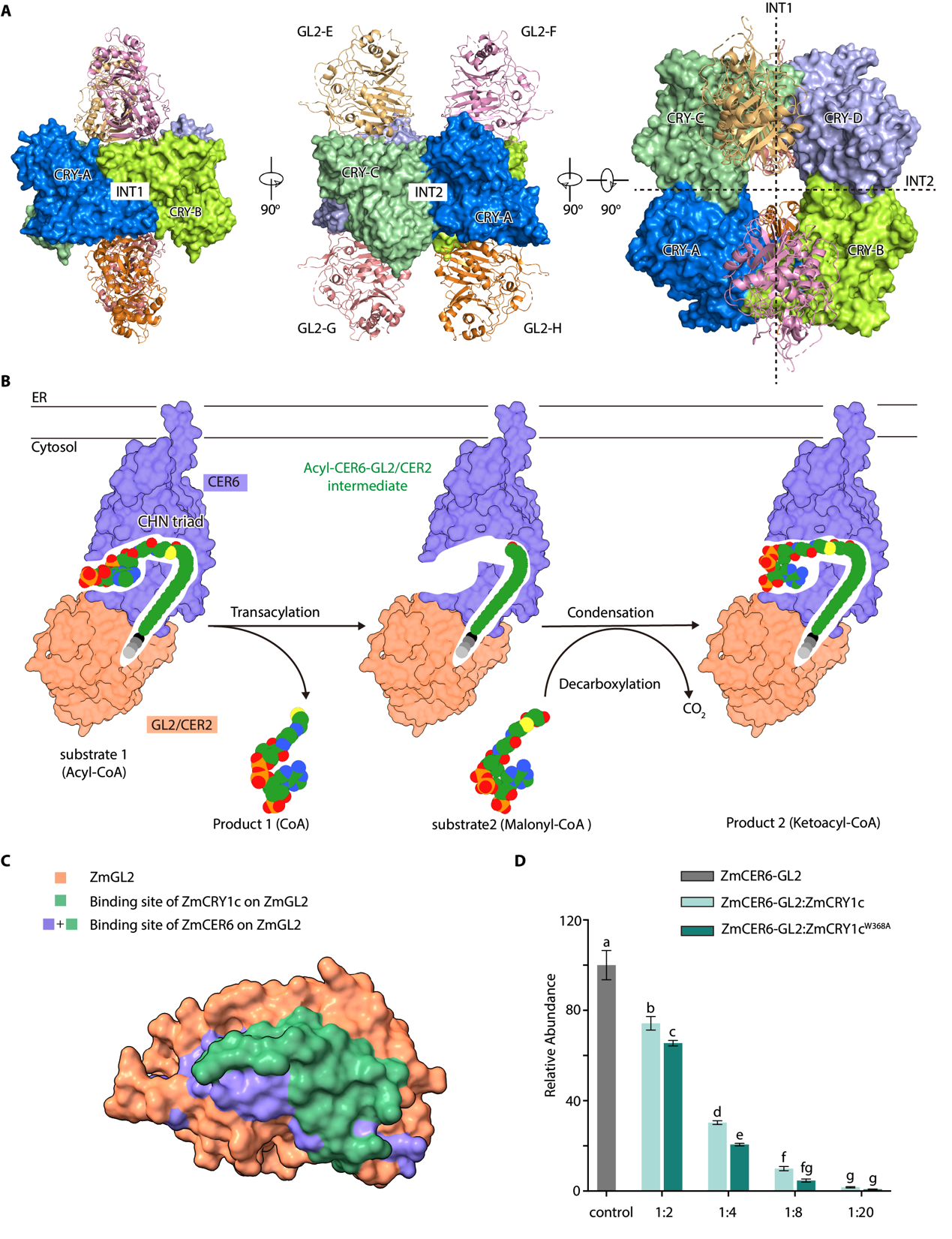
(A) ZmCRY1c-ZmGL2复合体三维结构。(B) ZmCER6-ZmGL2复合体催化超长链脂肪酸延伸的分子过程。(C-D) ZmCRY1c与ZmCER6竞争结合ZmGL2,调控ZmCER6-ZmGL2活性
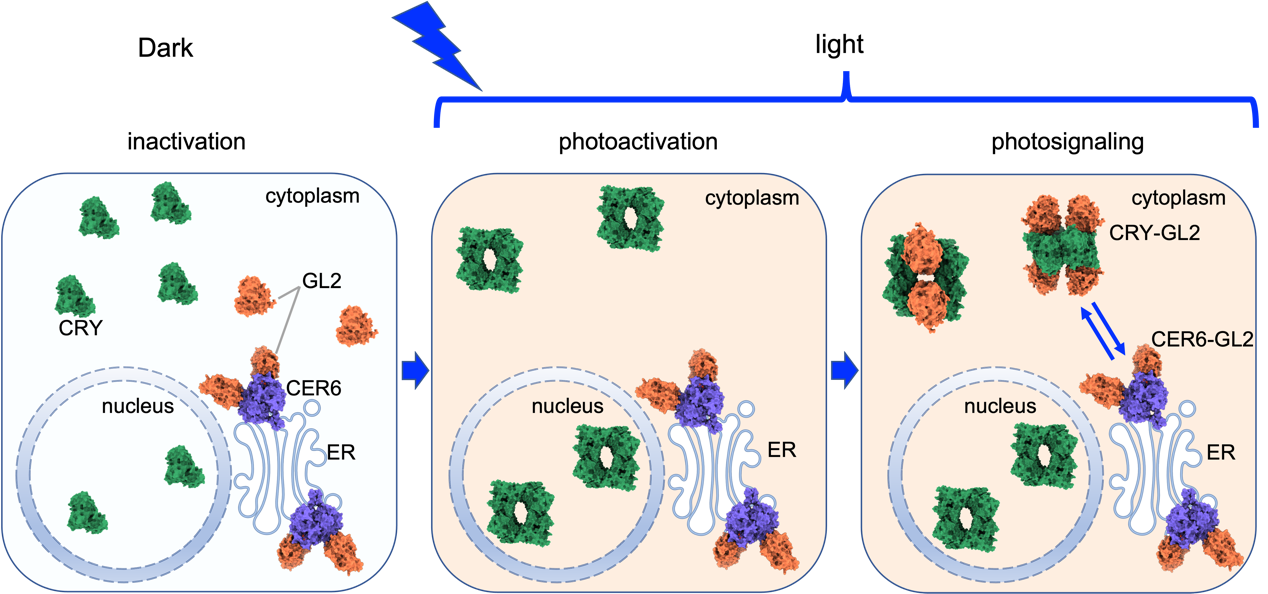
图2. 植物蓝光受体CRY信号复合体组装、调控蜡质合成的模型

Copyright © 2002-
中国科学院分子植物科学卓越创新中心 版权所有
地址:中国上海枫林路300号(200032)
电话:86-21-54924000
传真:86-21-54924015
Email: webmaster@cemps.ac.cn