

Chanhong Kim研究组发现光系统 II修复的氧化调控新机制
2026年6月14日,New Phytologist期刊在线发表了中国科学院分子植物科学卓越创新中心Chanhong Kim研究组题为 “Trp521 oxidation affects FtsH2 stability and its role in PSII repair”的研究论文。研究首次在叶绿体 FtsH2 蛋白酶上锁定一个氧化敏感色氨酸位点 Trp521,证实该位点是氧化胁迫下调控 PSII修复、维持叶绿体蛋白稳态的关键开关。
光合作用过程中伴随产生的光氧化胁迫时刻威胁叶绿体正常运转,强光、高温等环境剧烈波动时伤害更为突出。胁迫环境下,光系统 II(PSII)核心蛋白易遭受活性氧(ROS)氧化损伤;想要稳定光合效率、维持叶绿体内部稳态,受损蛋白必须快速降解并更新替换。承担这一修复降解工作的是叶绿体膜定位 FtsH 蛋白酶复合体,由 FtsH1、FtsH2、FtsH5、FtsH8 四种亚基组装而成。FtsH 复合体是 PSII蛋白质控的核心执行者,但长期以来,氧化胁迫究竟如何直接调控 FtsH 蛋白酶的稳定性与酶活,其中分子机制仍然未知。
色氨酸氧化修饰介导 PSII蛋白质控全新分子机制
研究发现,位于 FtsH2 蛋白水解结构域内的 Trp521 极易发生氧化修饰。研究人员结合质谱检测与转基因植株功能验证证实:Trp521 发生氧化后会破坏 FtsH2 蛋白稳定性,进而干扰光氧化胁迫下 D1、D2 等 PSII核心受损蛋白的降解周转进程。
关键位点突变实验直观印证该位点功能:将 Trp521 突变为ROS不敏感的亮氨酸(突变体 FtsH2W521L),FtsH2 蛋白稳定性大幅提升,植株在氧化胁迫环境下长势显著变好;反之,用苯丙氨酸替换(突变体 FtsH2W521F,模拟持续氧化修饰状态),会加速 FtsH2 自身降解,严重抑制植株生长。
研究进一步证实,当FtsH2的功能互补同源蛋白 FtsH8 缺失时,Trp521 位点突变带来的生长缺陷会急剧放大。这一结果充分说明,氧化修饰调控对维持 FtsH 六聚体完整结构、保障叶绿体蛋白质稳态不可或缺。
氧化敏感性是植物内源调控的开关
该研究表明FtsH2 的氧化修饰并非是单纯受 ROS 破坏的被动后果,而是植物自带的内源调控机制,可随外界环境变化动态 PSII的修复速率。
极具应用潜力的发现是:抗氧化型突变体 FtsH2W521L 可显著提升植株抗逆能力与生长表现。这意味着定向改造叶绿体蛋白、增强其抗氧化能力,有望成为提升作物光合效率、培育抗逆品种的优质育种思路。
与此同时,Trp521 位点在进化中高度保守,说明这种氧化敏感特性在特定自然环境中能赋予植物生存适应优势,也抛出了全新科学问题:植物如何在逆境中平衡蛋白结构稳定与调控灵活性?
这项成果清晰阐明氧化修饰如何调控叶绿体蛋白稳态与 PSII修复体系,拓展了学界对 “光合内源活性氧动态调控叶绿体蛋白” 这一过程的认知边界。此次 FtsH2 氧化调控位点的挖掘,为定向创制耐强光、耐光氧化胁迫、适应多变气候环境的高产抗逆作物打开全新育种靶点。
解析叶绿体蛋白酶应对氧化胁迫的应答机制是提升光合抗逆性的核心前提,该项研究证明 FtsH2 氧化修饰直接决定叶绿体蛋白质控水平,为培育高抗逆作物提供了崭新改造方向。同时,研究表明氧化调控是环境胁迫下调控叶绿体蛋白稳态、实现光合适应性的重要机制。
中国科学院分子植物科学卓越创新中心已毕业博士生张景志及Keun Pyo Lee副研究员为共同第一作者,Chanhong Kim研究员为通讯作者。凌祺桦研究组的博士生楼宇杰、Chanhong Kim研究组工作人员刘彦苓及凌祺桦研究员也参与了这项研究。本研究获得国家自然科学基金和植物性状形成与塑造全国重点实验室资助。
文章链接:http://doi.org/10.1111/nph.71356
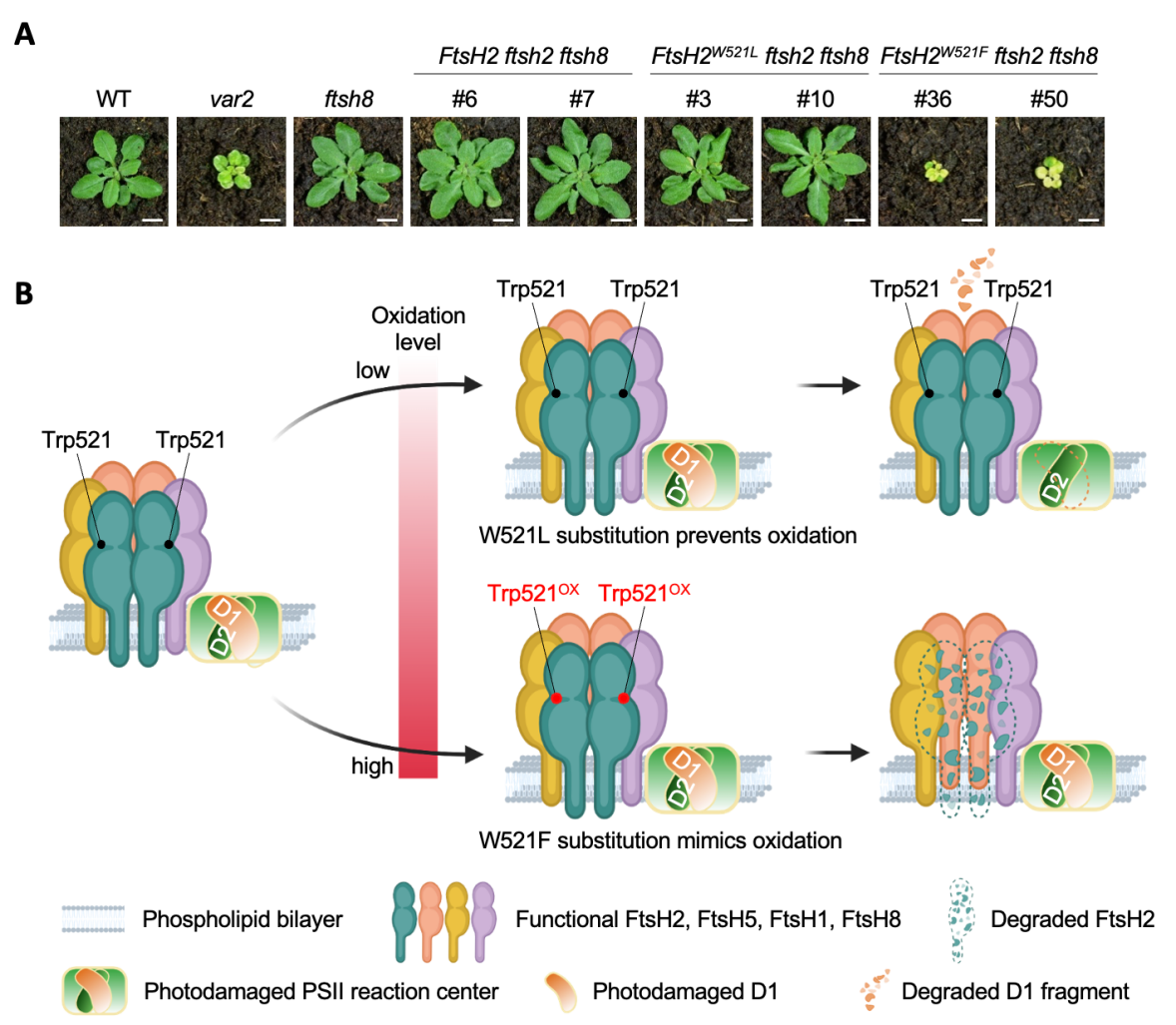
(A)B型FtsH缺失背景下FtsH2 Trp521不同类型点突的植株表型
(B)Trp521氧化修饰调控FtsH2稳定性与D1蛋白周转的工作模型

Copyright © 2002-
中国科学院分子植物科学卓越创新中心 版权所有
地址:中国上海枫林路300号(200032)
电话:86-21-54924000
传真:86-21-54924015
Email: webmaster@cemps.ac.cn